
Por recuperar mi juventud haría cualquier cosa en el mundo, salvo hacer ejercicio, madrugar, o ser respetable.
Oscar Wilde, El retrato de Dorian Gray
 |
| Detalle de Anciana y niño con velas (c. 1616–1617) de Pedro Pablo Rubens. |
LA ETERNA JUVENTUD es una de las aspiraciones perpetuas de la humanidad. Nadie es inmune a los efectos de la vejez, ya sea en nosotros mismos o en nuestros seres queridos. Sin embargo, más que un elemento ineludible de la condición humana, el envejecimiento es, de hecho, una característica biológica universal de los animales complejos, y quizá incluso de todos los seres vivos. Desde el punto de vista biológico, el envejecimiento es una disminución gradual de la capacidad de las células y tejidos del cuerpo para preservar su propia integridad y desempeñar sus funciones fisiológicas esenciales. La consecuencia última de este proceso es la incapacidad del cuerpo para sostener su propia existencia, conduciendo a una inevitable ‘muerte por vejez’. Sin importar cuánto esfuerzo se dedique a prolongar la vida, los seres humanos y otros animales parecen venir con una ‘fecha de caducidad’ intrínseca. Pero, ¿por qué ha de ser esto así? ¿Cuál es el origen de tan implacable fuerza de degeneración, y cómo es posible que los humanos seamos incapaces de derrotarla?
La cuestión de cuáles son las causas del envejecimiento, que se remonta a los días de Aristóteles, está en realidad compuesta por dos preguntas muy diferentes. La primera es la pregunta de por qué envejecemos: ¿cuál es la razón biológica última por la que los animales no han desarrollado la capacidad de vivir para siempre? La segunda pregunta es aquélla de cómo envejecemos: ¿cuáles son los procesos fisiológicos inmediatos que hacen que el cuerpo animal se deteriore con el tiempo? Aunque debería ser esperable que nuestro grado de conocimiento varíe entre estos dos niveles de análisis, quizá resulte sorprendente el que sea nuestra comprensión de cómo envejecemos, y no por qué envejecemos, la que actualmente se encuentra menos avanzada. En este ensayo se resume la perspectiva científica actual con respecto a estas dos dimensiones del proceso de envejecimiento.
Por qué envejecemos: las causas evolutivas del envejecimiento
La universalidad del envejecimiento en especies animales fue un hecho problemático para los primeros biólogos evolutivos. A mediados del siglo XIX, Charles Darwin propuso que los rasgos biológicos de las especies son producto de la evolución por selección natural y, por tanto, probablemente han sido útiles para la supervivencia y reproducción de generaciones pasadas. ¿Cómo es posible, entonces, que la evolución no haya producido organismos con la habilidad, claramente beneficiosa, de preservar su juventud indefinidamente?
La primera explicación evolutiva del envejecimiento fue propuesta por el biólogo August Weismann a finales del siglo XIX. Defensor temprano de las nuevas ideas de Darwin, Weismann fue una figura clave en el desarrollo de las primeras teorías sobre la herencia biológica. Para él, la paradoja evolutiva del envejecimiento podía resolverse a base de asumir que la longevidad de un animal es, en efecto, producto de la selección natural, pero no debido a un beneficio para el animal en sí, sino para la especie en su conjunto. Weismann propuso que la esperanza de vida de una especie ha evolucionado hasta un valor óptimo, el cual previene que la población se vea asfixiada por una preponderancia de individuos ancianos. Por tanto, según esta teoría, el envejecimiento es un mecanismo de mortalidad desarrollado específicamente para purgar a las generaciones más viejas y menos competitivas de la población, permitiendo así el éxito de individuos más jóvenes. Un detalle fascinante de esta teoría es su sorprendente coincidencia con las ideas del poeta y filósofo romano Lucrecio.
La explicación del envejecimiento propuesta por Weismann, pese a ser intuitivamente convincente, ha sido desmentida por los biólogos evolutivos de generaciones posteriores. Por una parte, el argumento de que los individuos ancianos deberían ser eliminados por ser menos competitivos que los individuos jóvenes invoca inmediatamente la suposición de que los animales experimentan un envejecimiento fisiológico. Sin embargo, para inferir los orígenes evolutivos del envejecimiento, es necesario partir de una población hipotética cuyos individuos no envejezcan y, por tanto, sólo puedan morir a causa de fuerzas extrínsecas como la depredación, la infección, el hambre o los accidentes. En dicha población, no hay razón para suponer que los individuos de mayor edad estarán en desventaja; en todo caso, el hecho de que hayan sobrevivido durante más tiempo implica que, en promedio, son mejores supervivientes. Además, los individuos de mayor edad contarán con una valiosa experiencia en lo que respecta a las tácticas y maniobras de la vida, de modo que deberían ofrecer una competencia formidable a los individuos jóvenes. Por tanto, sin la suposición previa de un proceso de envejecimiento, la muerte de los individuos ancianos no puede defenderse fácilmente como beneficiosa para la especie.
Otro poderoso argumento contra la teoría de Weismann es el hecho, ahora establecido, de que los rasgos que benefician al colectivo a expensas del individuo son evolutivamente inestables. En la mayoría de las situaciones, la selección natural opera abrumadoramente a nivel del individuo: si un ciervo, por ejemplo, es capaz de correr más rápido que sus congéneres, tendrá menor riesgo de ser depredado y, por lo tanto, mayor probabilidad de dejar descendencia, la cual heredará su superior velocidad. De la misma manera, si una especie desarrolla un proceso de envejecimiento que sea beneficioso para la especie pero perjudicial para el individuo, cualquier individuo que envejezca más lentamente que el resto tendrá una ventaja considerable —igual que el ciervo que es capaz de correr más rápido—, por lo que este rasgo se verá favorecido por la selección natural. El envejecimiento, por tanto, no puede haber evolucionado en beneficio exclusivo de la especie; si bien Weismann parece haber juzgado mal las implicaciones de la teoría de Darwin, podría alegarse en su defensa que al propio Darwin no le habría ido mejor. Ha sido sólo tras un siglo y medio de pensamiento que hemos llegado a entender el envejecimiento no como una consecuencia de la acción directa de la selección natural, sino más bien de su fracaso.
Una de las primeras versiones del concepto que subyace a las teorías modernas del envejecimiento fue propuesta por el influyente genetista matemático J.B.S. Haldane. Durante una inspirada serie de conferencias en 1940, Haldane señaló de pasada que la selección natural debería tener poco poder para eliminar un rasgo deletéreo si dicho rasgo solamente se manifiesta tarde en la vida del individuo. Para entender por qué esto es así, consideremos el caso de interés para Haldane: la enfermedad de Huntington. Pese a sus efectos devastadores y fatales, esta condición degenerativa generalmente comienza a manifestarse pasados los treinta años y, por tanto, tiene poco impacto en la capacidad de una persona para tener descendencia. Para cuando finalmente se diagnostica la enfermedad, es probable que los hijos del paciente ya hayan heredado el gen responsable. Haldane dedujo correctamente que éste es el motivo por el que la selección natural no ha sido capaz de suprimir un gen tan pernicioso. El impacto de la enfermedad de Huntington está confinado a la edad adulta, un periodo de la vida en el que la fuerza de la selección natural disminuye drásticamente, dado que la reproducción ya ha tenido lugar. Este periodo se denomina la ‘sombra selectiva’, porque los efectos biológicos confinados a esta etapa son prácticamente invisibles para la evolución.
El primero en aplicar el concepto de la sombra selectiva en la forma de una teoría completa del envejecimiento fue Peter Medawar, ganador del Premio Nobel en 1960. En la década de 1950, Medawar intentó explicar el envejecimiento como el efecto combinado de una colección de ‘genes mutantes’ —versiones alteradas de genes ‘normales’— cuyos efectos solamente aparecen relativamente tarde en la vida del individuo. Al igual que en el caso de la enfermedad de Huntington, las afecciones relacionadas con la edad, como las cataratas, la artritis y la osteoporosis, son de aparición tardía y no tienen impacto en la reproducción, lo cual impide que la selección natural elimine los genes mutantes implicados. Con el paso de miles de generaciones, un gran número de estos genes problemáticos se han ido acumulando ‘a la sombra’ de la selección, fusionándose sus efectos individuales para dar lugar a lo que llamamos envejecimiento. Medawar también captó la importancia de la mortalidad extrínseca, es decir, la tasa de muerte por fuerzas ambientales como la depredación: cuanto más tarde en la vida se expresen los efectos de un gen, menos individuos permanecerán vivos para experimentarlos. Por lo tanto, un gen que contribuya a prolongar la salud del músculo cardíaco durante muchas décadas podrá ser beneficioso para un elefante, pero carece de utilidad para un ratón que, con casi absoluta certeza, será depredado antes de cumplir los dos años.
Sobre la base del trabajo de Medawar, una teoría posterior propuso que el envejecimiento puede surgir de genes que no sólo tienen efectos negativos en la vejez, sino que también proporcionan beneficios en la juventud, cuando la selección natural tiene mayor fuerza. Según esta teoría, el envejecimiento sería un subproducto nocivo tardío de procesos que han sido favorecidos por ser beneficiosos en edades tempranas. El consenso científico actual es que cada una de estas teorías es probablemente correcta en ciertos casos, de forma que algunos componentes del envejecimiento se han originado a través de la acumulación de genes mutantes puramente perjudiciales, mientras que otros son efectos secundarios tardíos de genes beneficiosos.
Un aspecto importante de estas dos teorías evolutivas es que ambas definen el envejecimiento como el resultado de la incapacidad de la selección natural para mantener la integridad fisiológica durante más tiempo del que es realmente útil ‘en la naturaleza’. La idea fundamental es que no es ventajoso, evolutivamente hablando, vivir más de lo que ya vivimos, porque nuestra especie ha evolucionado para que podamos desarrollarnos y reproducirnos mucho antes de que nuestros cuerpos sucumban a la edad. Es más, debido a que el entorno natural de los primeros humanos hacía muy improbable que estos sobrevivieran tanto como nosotros lo hacemos, no ha habido ninguna necesidad evolutiva de una mayor longevidad. Hay que resaltar que nuestro modelo evolutivo del envejecimiento, el cual está bien respaldado por resultados teóricos y empíricos, no depende de qué mecanismos fisiológicos concretos sean responsables del envejecimiento. En otras palabras, aunque ciertamente entendemos por qué el proceso de envejecimiento existe en primer lugar, la escena es bastante distinta cuando consideramos la cuestión de cómo se desarrolla este proceso en un organismo dado.
Cómo envejecemos: causas mecánicas del envejecimiento
Afortunadamente para los científicos jóvenes, nuestras teorías mecanicistas del envejecimiento son mucho más abundantes y están peor respaldadas que las teorías evolutivas. Quizás la pregunta más inmediata con respecto al proceso de envejecimiento es si éste es consecuencia de un único mecanismo fisiológico o de múltiples mecanismos cuyos efectos están aproximadamente sincronizados. Dada la conclusión de que el envejecimiento es producto de la ineficacia de la selección natural, parece probable que este proceso debe de involucrar múltiples —posiblemente muchos— mecanismos no relacionados entre sí.
Como analogía rudimentaria, consideremos la situación de poseer un coche en una ciudad muy insegura, donde los vehículos son robados o dañados constantemente. En tales circunstancias, la decisión acertada sería adquirir un automóvil barato que pueda sobrevivir unos pocos años, y gastar lo menos posible en mantenimiento, ya que de lo contrario nuestra inversión bien podría ser un fracaso. No obstante, si por un golpe de suerte nos encontrásemos conduciendo el mismo coche al cabo de un buen número de años, no debería sorprendernos que nuestro vehículo nos decepcione en cualquier momento, debido precisamente a que es barato y está mal mantenido. Aunque esta analogía expone de manera poco halagadora la razón principal del envejecimiento —calidad y cuidado insuficientes—, no arroja luz alguna en lo que respecta a cuál de los componentes del coche se espera que falle primero. Dado que la degradación del coche es consecuencia de un mantenimiento deficiente, habríamos de esperar que muchos de sus componentes fallen con mayor y mayor frecuencia, hasta el punto en que la máquina en su conjunto sea incapaz de funcionar. Y diferentes procesos pueden ser responsables del fallo de distintos componentes: la transmisión podría desgastarse por pura fricción, mientras que los cilindros podrían sucumbir al hollín. Por lo tanto, aunque la causa última del envejecimiento pueda ser universal, los procesos inmediatamente involucrados en el mismo son múltiples y diversos.
Tal como sugiere esta analogía, la investigación actual sobre el envejecimiento se centra en la difícil tarea de establecer qué procesos fisiológicos contribuyen al envejecimiento, y cómo de importante es cada uno. Una gran variedad de procesos ha sido propuesta como causas mecánicas del envejecimiento; entre los más interesantes de estos se encuentran las ‘rutas de señalización de nutrientes’, que son redes funcionales de moléculas responsables de transmitir las señales fisiológicas que se generan cuando adquirimos nutrientes. La molécula más popular de esta red es la insulina, esencial para la regulación de los niveles de glucosa en sangre. Sin embargo, además de la bien conocida relación entre las deficiencias en la señalización de insulina y la diabetes, se ha descubierto que intervenciones biológicas que interfieren con la señalización de nutrientes pueden prolongar considerablemente la esperanza de vida de muchas especies, tanto vertebradas como invertebradas. Por ejemplo, un tratamiento conocido como ‘restricción calórica’, el cual consiste en limitar permanentemente el suministro de alimentos (o de ciertos nutrientes), se considera la forma más fiable de extender la vida en animales. Además, la desactivación de ciertos genes de señalización de nutrientes, ya sea por mutación o por tratamiento farmacológico, produce efectos similares a los de la restricción calórica. En la década de 1990, Cynthia Kenyon y sus compañeros descubrieron que mutaciones en uno de estos genes duplican la esperanza de vida de los gusanos nematodos, un hallazgo seguido de resultados similares en moscas de la fruta por los grupos de Linda Partridge y Marc Tatar. Por otra parte, la señalización de nutrientes también regula el crecimiento y desarrollo corporales, de modo que los animales sometidos a estas intervenciones tienden a estar atrofiados y mal desarrollados. Curiosamente, aunque la red de efectos moleculares mediante la cual la señalización de nutrientes modula el desarrollo y la longevidad aún no está completamente caracterizada, se cree que ésta es la razón de que las razas de perro pequeñas sean más longevas que las grandes.
Otro importante candidato entre los posibles mecanismos del envejecimiento es el daño molecular. Las células del cuerpo están constantemente expuestas a muchos tipos de daño químico, que pueden alterar las moléculas que las constituyen y comprometer la eficiencia de los procesos celulares. Los tipos de moléculas sujetas a este daño incluyen las proteínas (las cuales son tanto los ‘materiales de construcción’ de la célula como sus ‘herramientas de trabajo’) y el ADN (el cual almacena la información genética del organismo, incluidas las instrucciones para sintetizar proteínas). Un tipo de modificación del ADN que podría jugar un papel en el envejecimiento es el acortamiento de los telómeros, largos tramos de ADN que se encuentran en los extremos de los cromosomas para preservar su estructura, como el herrete al final del cordón de un zapato. Los telómeros se acortan ligeramente cada vez que una célula se divide en dos, hasta que, finalmente, se vuelven demasiado cortos para permitir nuevas divisiones celulares. Aunque se cree que esta erosión de los telómeros constituye una barrera importante contra el cáncer, es posible que también sea una causa del envejecimiento. Recientemente, la bióloga María Blasco y su equipo informaron del sorprendente hallazgo de que la tasa de acortamiento de los telómeros en una especie está relacionada con su esperanza de vida, de modo que los telómeros se erosionan más rápido en especies de vida más corta. No obstante, esta relación se ve oscurecida por el hecho de que las especies con menor esperanza de vida también tienden a ser más pequeñas, y se sabe que el tamaño corporal influye en muchos aspectos de la fisiología animal.
La cuestión de cuáles son las causas del envejecimiento, que se remonta a los días de Aristóteles, está en realidad compuesta por dos preguntas muy diferentes. La primera es la pregunta de por qué envejecemos: ¿cuál es la razón biológica última por la que los animales no han desarrollado la capacidad de vivir para siempre? La segunda pregunta es aquélla de cómo envejecemos: ¿cuáles son los procesos fisiológicos inmediatos que hacen que el cuerpo animal se deteriore con el tiempo? Aunque debería ser esperable que nuestro grado de conocimiento varíe entre estos dos niveles de análisis, quizá resulte sorprendente el que sea nuestra comprensión de cómo envejecemos, y no por qué envejecemos, la que actualmente se encuentra menos avanzada. En este ensayo se resume la perspectiva científica actual con respecto a estas dos dimensiones del proceso de envejecimiento.
Por qué envejecemos: las causas evolutivas del envejecimiento
La universalidad del envejecimiento en especies animales fue un hecho problemático para los primeros biólogos evolutivos. A mediados del siglo XIX, Charles Darwin propuso que los rasgos biológicos de las especies son producto de la evolución por selección natural y, por tanto, probablemente han sido útiles para la supervivencia y reproducción de generaciones pasadas. ¿Cómo es posible, entonces, que la evolución no haya producido organismos con la habilidad, claramente beneficiosa, de preservar su juventud indefinidamente?
La primera explicación evolutiva del envejecimiento fue propuesta por el biólogo August Weismann a finales del siglo XIX. Defensor temprano de las nuevas ideas de Darwin, Weismann fue una figura clave en el desarrollo de las primeras teorías sobre la herencia biológica. Para él, la paradoja evolutiva del envejecimiento podía resolverse a base de asumir que la longevidad de un animal es, en efecto, producto de la selección natural, pero no debido a un beneficio para el animal en sí, sino para la especie en su conjunto. Weismann propuso que la esperanza de vida de una especie ha evolucionado hasta un valor óptimo, el cual previene que la población se vea asfixiada por una preponderancia de individuos ancianos. Por tanto, según esta teoría, el envejecimiento es un mecanismo de mortalidad desarrollado específicamente para purgar a las generaciones más viejas y menos competitivas de la población, permitiendo así el éxito de individuos más jóvenes. Un detalle fascinante de esta teoría es su sorprendente coincidencia con las ideas del poeta y filósofo romano Lucrecio.
La explicación del envejecimiento propuesta por Weismann, pese a ser intuitivamente convincente, ha sido desmentida por los biólogos evolutivos de generaciones posteriores. Por una parte, el argumento de que los individuos ancianos deberían ser eliminados por ser menos competitivos que los individuos jóvenes invoca inmediatamente la suposición de que los animales experimentan un envejecimiento fisiológico. Sin embargo, para inferir los orígenes evolutivos del envejecimiento, es necesario partir de una población hipotética cuyos individuos no envejezcan y, por tanto, sólo puedan morir a causa de fuerzas extrínsecas como la depredación, la infección, el hambre o los accidentes. En dicha población, no hay razón para suponer que los individuos de mayor edad estarán en desventaja; en todo caso, el hecho de que hayan sobrevivido durante más tiempo implica que, en promedio, son mejores supervivientes. Además, los individuos de mayor edad contarán con una valiosa experiencia en lo que respecta a las tácticas y maniobras de la vida, de modo que deberían ofrecer una competencia formidable a los individuos jóvenes. Por tanto, sin la suposición previa de un proceso de envejecimiento, la muerte de los individuos ancianos no puede defenderse fácilmente como beneficiosa para la especie.
Otro poderoso argumento contra la teoría de Weismann es el hecho, ahora establecido, de que los rasgos que benefician al colectivo a expensas del individuo son evolutivamente inestables. En la mayoría de las situaciones, la selección natural opera abrumadoramente a nivel del individuo: si un ciervo, por ejemplo, es capaz de correr más rápido que sus congéneres, tendrá menor riesgo de ser depredado y, por lo tanto, mayor probabilidad de dejar descendencia, la cual heredará su superior velocidad. De la misma manera, si una especie desarrolla un proceso de envejecimiento que sea beneficioso para la especie pero perjudicial para el individuo, cualquier individuo que envejezca más lentamente que el resto tendrá una ventaja considerable —igual que el ciervo que es capaz de correr más rápido—, por lo que este rasgo se verá favorecido por la selección natural. El envejecimiento, por tanto, no puede haber evolucionado en beneficio exclusivo de la especie; si bien Weismann parece haber juzgado mal las implicaciones de la teoría de Darwin, podría alegarse en su defensa que al propio Darwin no le habría ido mejor. Ha sido sólo tras un siglo y medio de pensamiento que hemos llegado a entender el envejecimiento no como una consecuencia de la acción directa de la selección natural, sino más bien de su fracaso.
Una de las primeras versiones del concepto que subyace a las teorías modernas del envejecimiento fue propuesta por el influyente genetista matemático J.B.S. Haldane. Durante una inspirada serie de conferencias en 1940, Haldane señaló de pasada que la selección natural debería tener poco poder para eliminar un rasgo deletéreo si dicho rasgo solamente se manifiesta tarde en la vida del individuo. Para entender por qué esto es así, consideremos el caso de interés para Haldane: la enfermedad de Huntington. Pese a sus efectos devastadores y fatales, esta condición degenerativa generalmente comienza a manifestarse pasados los treinta años y, por tanto, tiene poco impacto en la capacidad de una persona para tener descendencia. Para cuando finalmente se diagnostica la enfermedad, es probable que los hijos del paciente ya hayan heredado el gen responsable. Haldane dedujo correctamente que éste es el motivo por el que la selección natural no ha sido capaz de suprimir un gen tan pernicioso. El impacto de la enfermedad de Huntington está confinado a la edad adulta, un periodo de la vida en el que la fuerza de la selección natural disminuye drásticamente, dado que la reproducción ya ha tenido lugar. Este periodo se denomina la ‘sombra selectiva’, porque los efectos biológicos confinados a esta etapa son prácticamente invisibles para la evolución.
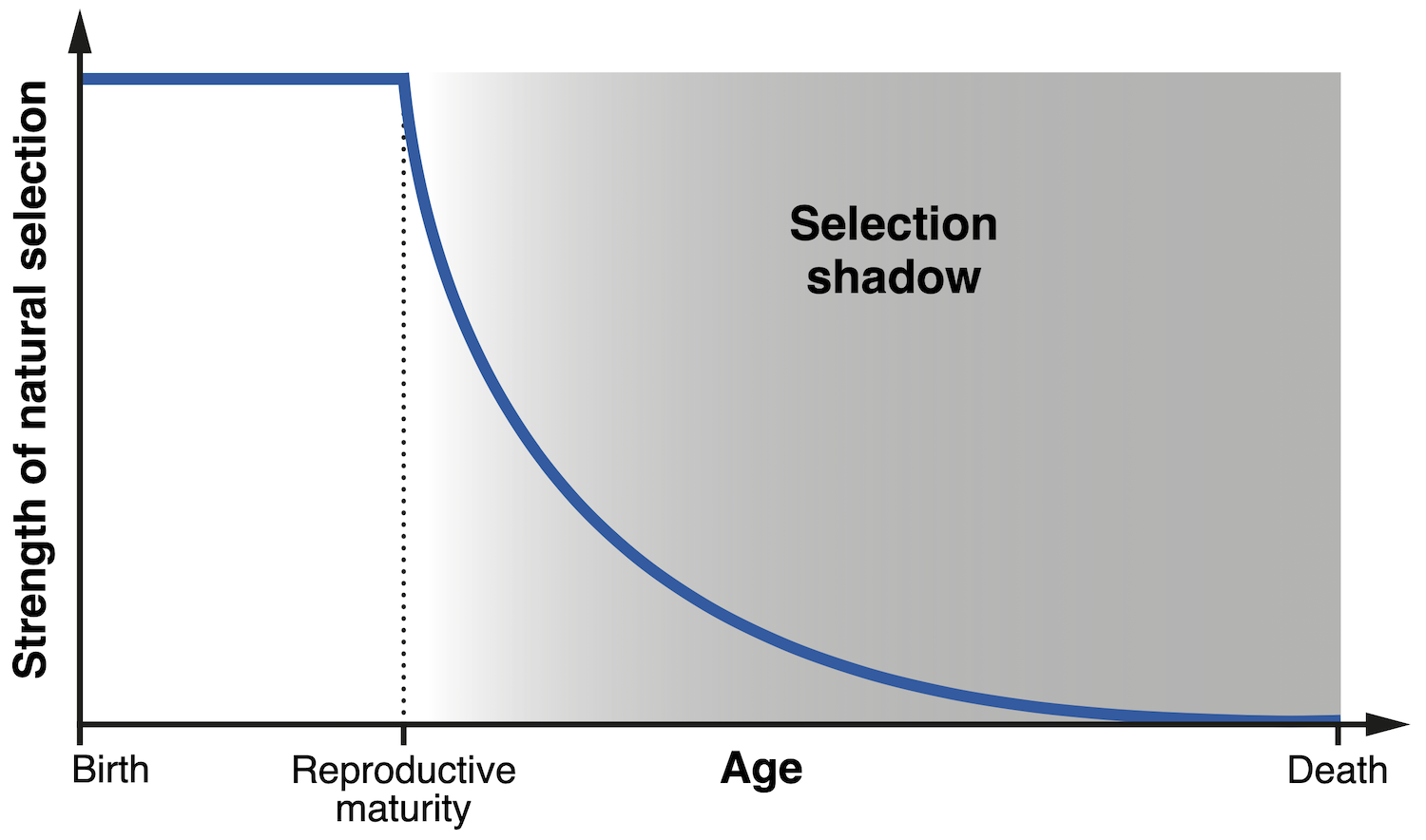 |
| Diagrama que ilustra el concepto de ‘sombra selectiva’ (selection shadow), que se refiere a la disminución progresiva de la fuerza de la selección natural pasada la edad de madurez reproductiva (A. Báez Ortega). |
El primero en aplicar el concepto de la sombra selectiva en la forma de una teoría completa del envejecimiento fue Peter Medawar, ganador del Premio Nobel en 1960. En la década de 1950, Medawar intentó explicar el envejecimiento como el efecto combinado de una colección de ‘genes mutantes’ —versiones alteradas de genes ‘normales’— cuyos efectos solamente aparecen relativamente tarde en la vida del individuo. Al igual que en el caso de la enfermedad de Huntington, las afecciones relacionadas con la edad, como las cataratas, la artritis y la osteoporosis, son de aparición tardía y no tienen impacto en la reproducción, lo cual impide que la selección natural elimine los genes mutantes implicados. Con el paso de miles de generaciones, un gran número de estos genes problemáticos se han ido acumulando ‘a la sombra’ de la selección, fusionándose sus efectos individuales para dar lugar a lo que llamamos envejecimiento. Medawar también captó la importancia de la mortalidad extrínseca, es decir, la tasa de muerte por fuerzas ambientales como la depredación: cuanto más tarde en la vida se expresen los efectos de un gen, menos individuos permanecerán vivos para experimentarlos. Por lo tanto, un gen que contribuya a prolongar la salud del músculo cardíaco durante muchas décadas podrá ser beneficioso para un elefante, pero carece de utilidad para un ratón que, con casi absoluta certeza, será depredado antes de cumplir los dos años.
Sobre la base del trabajo de Medawar, una teoría posterior propuso que el envejecimiento puede surgir de genes que no sólo tienen efectos negativos en la vejez, sino que también proporcionan beneficios en la juventud, cuando la selección natural tiene mayor fuerza. Según esta teoría, el envejecimiento sería un subproducto nocivo tardío de procesos que han sido favorecidos por ser beneficiosos en edades tempranas. El consenso científico actual es que cada una de estas teorías es probablemente correcta en ciertos casos, de forma que algunos componentes del envejecimiento se han originado a través de la acumulación de genes mutantes puramente perjudiciales, mientras que otros son efectos secundarios tardíos de genes beneficiosos.
Un aspecto importante de estas dos teorías evolutivas es que ambas definen el envejecimiento como el resultado de la incapacidad de la selección natural para mantener la integridad fisiológica durante más tiempo del que es realmente útil ‘en la naturaleza’. La idea fundamental es que no es ventajoso, evolutivamente hablando, vivir más de lo que ya vivimos, porque nuestra especie ha evolucionado para que podamos desarrollarnos y reproducirnos mucho antes de que nuestros cuerpos sucumban a la edad. Es más, debido a que el entorno natural de los primeros humanos hacía muy improbable que estos sobrevivieran tanto como nosotros lo hacemos, no ha habido ninguna necesidad evolutiva de una mayor longevidad. Hay que resaltar que nuestro modelo evolutivo del envejecimiento, el cual está bien respaldado por resultados teóricos y empíricos, no depende de qué mecanismos fisiológicos concretos sean responsables del envejecimiento. En otras palabras, aunque ciertamente entendemos por qué el proceso de envejecimiento existe en primer lugar, la escena es bastante distinta cuando consideramos la cuestión de cómo se desarrolla este proceso en un organismo dado.
Cómo envejecemos: causas mecánicas del envejecimiento
Afortunadamente para los científicos jóvenes, nuestras teorías mecanicistas del envejecimiento son mucho más abundantes y están peor respaldadas que las teorías evolutivas. Quizás la pregunta más inmediata con respecto al proceso de envejecimiento es si éste es consecuencia de un único mecanismo fisiológico o de múltiples mecanismos cuyos efectos están aproximadamente sincronizados. Dada la conclusión de que el envejecimiento es producto de la ineficacia de la selección natural, parece probable que este proceso debe de involucrar múltiples —posiblemente muchos— mecanismos no relacionados entre sí.
Como analogía rudimentaria, consideremos la situación de poseer un coche en una ciudad muy insegura, donde los vehículos son robados o dañados constantemente. En tales circunstancias, la decisión acertada sería adquirir un automóvil barato que pueda sobrevivir unos pocos años, y gastar lo menos posible en mantenimiento, ya que de lo contrario nuestra inversión bien podría ser un fracaso. No obstante, si por un golpe de suerte nos encontrásemos conduciendo el mismo coche al cabo de un buen número de años, no debería sorprendernos que nuestro vehículo nos decepcione en cualquier momento, debido precisamente a que es barato y está mal mantenido. Aunque esta analogía expone de manera poco halagadora la razón principal del envejecimiento —calidad y cuidado insuficientes—, no arroja luz alguna en lo que respecta a cuál de los componentes del coche se espera que falle primero. Dado que la degradación del coche es consecuencia de un mantenimiento deficiente, habríamos de esperar que muchos de sus componentes fallen con mayor y mayor frecuencia, hasta el punto en que la máquina en su conjunto sea incapaz de funcionar. Y diferentes procesos pueden ser responsables del fallo de distintos componentes: la transmisión podría desgastarse por pura fricción, mientras que los cilindros podrían sucumbir al hollín. Por lo tanto, aunque la causa última del envejecimiento pueda ser universal, los procesos inmediatamente involucrados en el mismo son múltiples y diversos.
Tal como sugiere esta analogía, la investigación actual sobre el envejecimiento se centra en la difícil tarea de establecer qué procesos fisiológicos contribuyen al envejecimiento, y cómo de importante es cada uno. Una gran variedad de procesos ha sido propuesta como causas mecánicas del envejecimiento; entre los más interesantes de estos se encuentran las ‘rutas de señalización de nutrientes’, que son redes funcionales de moléculas responsables de transmitir las señales fisiológicas que se generan cuando adquirimos nutrientes. La molécula más popular de esta red es la insulina, esencial para la regulación de los niveles de glucosa en sangre. Sin embargo, además de la bien conocida relación entre las deficiencias en la señalización de insulina y la diabetes, se ha descubierto que intervenciones biológicas que interfieren con la señalización de nutrientes pueden prolongar considerablemente la esperanza de vida de muchas especies, tanto vertebradas como invertebradas. Por ejemplo, un tratamiento conocido como ‘restricción calórica’, el cual consiste en limitar permanentemente el suministro de alimentos (o de ciertos nutrientes), se considera la forma más fiable de extender la vida en animales. Además, la desactivación de ciertos genes de señalización de nutrientes, ya sea por mutación o por tratamiento farmacológico, produce efectos similares a los de la restricción calórica. En la década de 1990, Cynthia Kenyon y sus compañeros descubrieron que mutaciones en uno de estos genes duplican la esperanza de vida de los gusanos nematodos, un hallazgo seguido de resultados similares en moscas de la fruta por los grupos de Linda Partridge y Marc Tatar. Por otra parte, la señalización de nutrientes también regula el crecimiento y desarrollo corporales, de modo que los animales sometidos a estas intervenciones tienden a estar atrofiados y mal desarrollados. Curiosamente, aunque la red de efectos moleculares mediante la cual la señalización de nutrientes modula el desarrollo y la longevidad aún no está completamente caracterizada, se cree que ésta es la razón de que las razas de perro pequeñas sean más longevas que las grandes.
Otro importante candidato entre los posibles mecanismos del envejecimiento es el daño molecular. Las células del cuerpo están constantemente expuestas a muchos tipos de daño químico, que pueden alterar las moléculas que las constituyen y comprometer la eficiencia de los procesos celulares. Los tipos de moléculas sujetas a este daño incluyen las proteínas (las cuales son tanto los ‘materiales de construcción’ de la célula como sus ‘herramientas de trabajo’) y el ADN (el cual almacena la información genética del organismo, incluidas las instrucciones para sintetizar proteínas). Un tipo de modificación del ADN que podría jugar un papel en el envejecimiento es el acortamiento de los telómeros, largos tramos de ADN que se encuentran en los extremos de los cromosomas para preservar su estructura, como el herrete al final del cordón de un zapato. Los telómeros se acortan ligeramente cada vez que una célula se divide en dos, hasta que, finalmente, se vuelven demasiado cortos para permitir nuevas divisiones celulares. Aunque se cree que esta erosión de los telómeros constituye una barrera importante contra el cáncer, es posible que también sea una causa del envejecimiento. Recientemente, la bióloga María Blasco y su equipo informaron del sorprendente hallazgo de que la tasa de acortamiento de los telómeros en una especie está relacionada con su esperanza de vida, de modo que los telómeros se erosionan más rápido en especies de vida más corta. No obstante, esta relación se ve oscurecida por el hecho de que las especies con menor esperanza de vida también tienden a ser más pequeñas, y se sabe que el tamaño corporal influye en muchos aspectos de la fisiología animal.
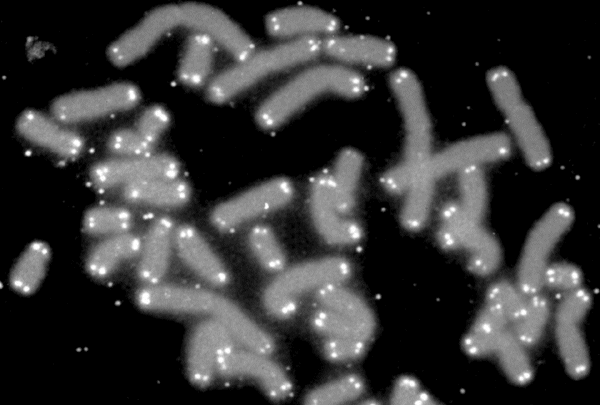 |
| Imagen de microscopía de fluorescencia que muestra la ubicación de los telómeros (en blanco) en los extremos de los cromosomas de una célula humana (en gris). Los telómeros preservan la integridad del ADN en cada cromosoma, y se ha propuesto que su acortamiento con el tiempo es una causa del envejecimiento (NASA/Wikimedia Commons, dominio público). |
Recientemente, trabajando junto con Alex Cagan, Íñigo Martincorena y otros investigadores del Wellcome Sanger Institute, hemos explorado la relación entre la esperanza de vida y otra forma común de modificación del ADN: las mutaciones somáticas. Este término se refiere a los cambios que se acumulan en nuestro ADN con el tiempo; tales mutaciones no están presentes inicialmente en ninguna de nuestras células, sino que van siendo adquiridas por células individuales a medida que nuestros cuerpos crecen y envejecen. La hipótesis de que las mutaciones somáticas contribuyen al envejecimiento se planteó por primera vez en la década de 1960, pero su papel exacto sigue siendo una incógnita. Tras caracterizar la tasa de mutación en dieciséis especies de mamíferos, desde ratones hasta jirafas, encontramos una relación muy similar a la descrita para los telómeros: las especies de vida corta mutan más rápido que las de vida más larga, de tal modo que una célula de ratón adquiere tantas mutaciones en dos años como una célula humana en ochenta. Concluimos, además, que este resultado no se ve afectado por la relación entre la longevidad y el tamaño corporal: al menos en mamíferos, la tasa de mutación somática puede emplearse para predecir la esperanza de vida de una especie, independientemente de su tamaño. El hecho de que las tasas de diferentes formas de daño molecular presentan relaciones similares con la esperanza de vida sugiere —aunque no demuestra— que estas clases de daño pueden estar involucradas en el envejecimiento.
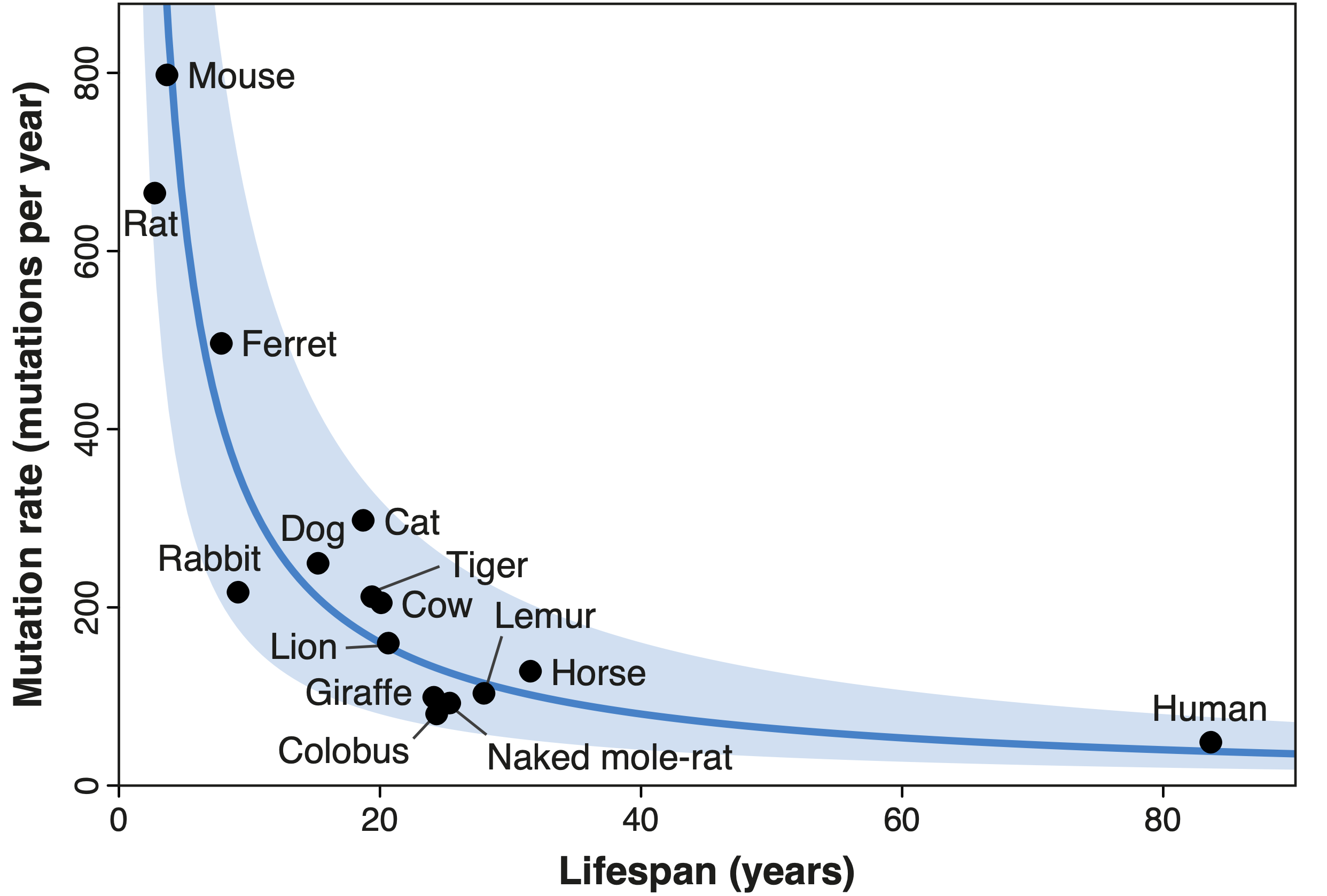 |
| Diagrama que muestra la relación inversa entre la esperanza de vida (lifespan) y la tasa de mutación somática (mutation rate) en 16 especies de mamíferos. La tasa de mutación de cada especie es inversamente proporcional a su esperanza de vida, tal que todas las especies tienen un número similar de mutaciones en su ADN al final de sus respectivas vidas. Esta relación está indicada por la línea azul, con el área sombreada marcando una desviación de esta línea por un factor de dos (Fuente: Cagan, Baez-Ortega et al., 2022). |
Aunque pueda parecer inconsistente que procesos tan dispares como la señalización de nutrientes y el daño molecular contribuyan al envejecimiento, estos procesos no son tan remotos cuando se observan a la luz de una teoría del envejecimiento conocida como la teoría del ‘soma desechable’. Según esta explicación, la fisiología de los organismos complejos incorpora un equilibrio energético central, mediante el cual la energía adquirida de los nutrientes se distribuye entre los procesos de mantenimiento somático (la preservación del cuerpo mediante la reparación del daño molecular) y reproducción (la preservación de los genes mediante su transmisión a la descendencia). En lugar de lidiar con el origen evolutivo del envejecimiento, esta teoría proporciona un marco para entender su regulación fisiológica. Dado que el cuerpo (o ‘soma’) es, en última instancia, perecedero, el equilibrio energético entre el mantenimiento y la reproducción supuestamente ha sido optimizado por la evolución para favorecer el costoso proceso de reproducción en tiempos de abundancia, y promover procesos de mantenimiento cuando hay escasez de nutrientes. Por tanto, es posible que disrupciones en la señalización de nutrientes modifiquen la tasa de envejecimiento por interferir con el ‘medidor’ de este sistema de asignación de energía, mientras que el daño molecular puede ser simplemente la fuerza que se opone a los procesos de mantenimiento somático. A pesar de la notable coherencia de la teoría del soma desechable, la evidencia de la existencia de un equilibrio energético universal en animales todavía no es concluyente. Es posible que, como tantas otras cosas en la biología, los sistemas de distribución de energía sean cruciales pero no universales: puede que sean relevantes sólo en ciertas especies, o en algunos órganos, o en periodos concretos de la vida. Incluso en esta época de progreso científico sin precedentes, existe una inmensidad de conocimiento por descubrir acerca de los procesos fisiológicos que contribuyen al envejecimiento.
La batalla contra el envejecimiento
Desde los días de Darwin y Weismann, hemos llegado a comprender el envejecimiento no como una ‘fuerza mortal’ dedicada al beneficio de la especie, sino como una consecuencia inevitable de la forma en que opera la evolución. Los cuerpos animales no han evolucionado con objeto de vivir para siempre, sino de sobrevivir y reproducirse en un entorno despiadado. Nuestra biología es tal y como es precisamente porque nuestros antepasados tuvieron éxito en estas metas, no porque consiguieron vivir para siempre.
Cualesquiera que sean las causas del envejecimiento, la pregunta fundamental para la humanidad es si alguna vez lograremos controlarlas, quizá no con miras a vivir para siempre, sino a disfrutar, al menos, de una salud más duradera y una vejez más feliz. Está claro que este objetivo habrá de permanecer fuera de nuestro alcance mientras no entendamos qué significa exactamente ‘envejecer’ a nivel molecular. Puede que algún día obtengamos el poder de manipular los procesos mediante los cuales nuestros cuerpos mantienen a raya los efectos del tiempo, o incluso de combatir dichos efectos directamente; puede que finalmente seamos capaces de someter y domesticar el proceso de envejecimiento. Pero tales milagros aguardan aún tras el horizonte; en años venideros, tendremos que seguir aprovechando la capacidad de la medicina moderna para tratar cada una de las aflicciones relacionadas con la edad.
Cuando se trata de hacerse viejo, la teoría personal de A.C. Benson —ensayista, poeta y antiguo director (Master) del Magdalene College de Cambridge— tal vez resulte más provechosa que las aquí discutidas: ‘Tengo la teoría de que uno ha de envejecer de forma tranquila y adecuada, que uno ha de estar perfectamente satisfecho con su época en la vida, que las diversiones y ocupaciones deben cambiar natural y fácilmente, y no ser abandonadas con pesadumbre’. Una teoría algo modesta, quizá; Benson no tarda en admitir que ‘es más fácil decir que hacer’. Sin embargo, aun cuando seamos conscientes de la lenta e impasible fuga de la juventud por entre nuestros dedos, conviene no olvidar las palabras de Longfellow:
La batalla contra el envejecimiento
Desde los días de Darwin y Weismann, hemos llegado a comprender el envejecimiento no como una ‘fuerza mortal’ dedicada al beneficio de la especie, sino como una consecuencia inevitable de la forma en que opera la evolución. Los cuerpos animales no han evolucionado con objeto de vivir para siempre, sino de sobrevivir y reproducirse en un entorno despiadado. Nuestra biología es tal y como es precisamente porque nuestros antepasados tuvieron éxito en estas metas, no porque consiguieron vivir para siempre.
Cualesquiera que sean las causas del envejecimiento, la pregunta fundamental para la humanidad es si alguna vez lograremos controlarlas, quizá no con miras a vivir para siempre, sino a disfrutar, al menos, de una salud más duradera y una vejez más feliz. Está claro que este objetivo habrá de permanecer fuera de nuestro alcance mientras no entendamos qué significa exactamente ‘envejecer’ a nivel molecular. Puede que algún día obtengamos el poder de manipular los procesos mediante los cuales nuestros cuerpos mantienen a raya los efectos del tiempo, o incluso de combatir dichos efectos directamente; puede que finalmente seamos capaces de someter y domesticar el proceso de envejecimiento. Pero tales milagros aguardan aún tras el horizonte; en años venideros, tendremos que seguir aprovechando la capacidad de la medicina moderna para tratar cada una de las aflicciones relacionadas con la edad.
Cuando se trata de hacerse viejo, la teoría personal de A.C. Benson —ensayista, poeta y antiguo director (Master) del Magdalene College de Cambridge— tal vez resulte más provechosa que las aquí discutidas: ‘Tengo la teoría de que uno ha de envejecer de forma tranquila y adecuada, que uno ha de estar perfectamente satisfecho con su época en la vida, que las diversiones y ocupaciones deben cambiar natural y fácilmente, y no ser abandonadas con pesadumbre’. Una teoría algo modesta, quizá; Benson no tarda en admitir que ‘es más fácil decir que hacer’. Sin embargo, aun cuando seamos conscientes de la lenta e impasible fuga de la juventud por entre nuestros dedos, conviene no olvidar las palabras de Longfellow:
Pues la vejez es tanto una oportunidad,
Con otro vestido, como la mocedad,
Y en el crepúsculo se viste el firmamento
De estrellas invisibles hasta ese momento.
Referencias
Weismann, A. ‘The duration of life’ (1881). In Essays Upon Heredity and Kindred Biological Problems (tr. Poulton, EB, Schönland, S, Shipley, AE). Clarendon, 1889.
Haldane, JBS. New Paths in Genetics. Allen & Unwin, 1941.
Kenyon, C, Chang, J et al. A C. elegans mutant that lives twice as long as wild type. Nature, 1993.
Hughes, KA, Reynolds, RM. Evolutionary and mechanistic theories of aging. Annual Review of Entomology, 2005.
Kirkwood, TBL. Understanding the odd science of aging. Cell, 2005.
Flatt, T, Partridge, L. Horizons in the evolution of aging. BMC Biology, 2018.
Whittemore, K, Vera, E et al. Telomere shortening rate predicts species life span. Proceedings of the National Academy of Sciences, 2019.
Cagan, A, Baez-Ortega, A et al. Somatic mutation rates scale with lifespan across mammals. Nature, 2022.
Este artículo es una traducción de un artículo publicado en el Magdalene College Magazine (2021–22).
El autor agradece a James Raven y Aude Fitzsimons sus comentarios sobre el manuscrito original.
Weismann, A. ‘The duration of life’ (1881). In Essays Upon Heredity and Kindred Biological Problems (tr. Poulton, EB, Schönland, S, Shipley, AE). Clarendon, 1889.
Haldane, JBS. New Paths in Genetics. Allen & Unwin, 1941.
Kenyon, C, Chang, J et al. A C. elegans mutant that lives twice as long as wild type. Nature, 1993.
Hughes, KA, Reynolds, RM. Evolutionary and mechanistic theories of aging. Annual Review of Entomology, 2005.
Kirkwood, TBL. Understanding the odd science of aging. Cell, 2005.
Flatt, T, Partridge, L. Horizons in the evolution of aging. BMC Biology, 2018.
Whittemore, K, Vera, E et al. Telomere shortening rate predicts species life span. Proceedings of the National Academy of Sciences, 2019.
Cagan, A, Baez-Ortega, A et al. Somatic mutation rates scale with lifespan across mammals. Nature, 2022.
Este artículo es una traducción de un artículo publicado en el Magdalene College Magazine (2021–22).
El autor agradece a James Raven y Aude Fitzsimons sus comentarios sobre el manuscrito original.